Bitki üremesi, bitkilerin yeni bireyler veya yavrular ürettikleri süreçtir. Üreme ya cinsel ya da eşeysizdir. Cinsel üreme, gametlerin füzyonu ile yavruların oluşmasıdır. . Eşeysiz üreme, gametlerin füzyonu olmadan yavruların oluşmasıdır. Cinsel üreme, yavruların genetik olarak ebeveynlerden farklı olmasına neden olur. Eşeysiz yavrular, mutasyon dışında genetik olarak özdeştir. Daha yüksek bitkilerde, yavrular, uzun ömürlü olabilen ve yavruları ebeveynlerden biraz uzaklara dağıtabilen koruyucu bir tohumda paketlenir. Çiçekli bitkilerde (kapalı tohumlular), tohumun kendisi, gelişmekte olan tohumları koruyabilen ve dağılmasına yardımcı olabilecek bir meyvenin içinde bulunur.

Kapalı Alanlarda Cinsel Üreme: Ovül Oluşumu
Tüm bitkilerin, boyut ve hücre başına kromozom sayısı bakımından farklılık gösteren iki farklı formdan oluşan bir yaşam döngüsü vardır . Çiçekli bitkilerde
Kökler, sürgünler, yapraklar ve üreme yapılarından (çiçekler ve meyve) oluşan büyük, tanıdık form diploiddir ve sporofit olarak adlandırılır. Sporofit , çiçek tarafından üretilen dokulara bağlı haploid mikroskobik gametofitleri üretir . Çiçekli bir bitkinin üreme döngüsü, sporofitten gametofit'e gidip gelen düzenli, genellikle mevsimseldir.
Çiçek, erkek ve dişi olmak üzere iki tür gametofit üretir. Dişi gametofit , çiçeğin yumurtalıkındaki küçük bir yapı olan ovül içindeki bir hücreden doğar . Yumurtalık, çiçeğin içinde genellikle birçok yumurtayı barındıran ve koruyan daha büyük bir yapıdır. Çiçekli bitkiler, yumurtalarının tamamen yumurtalık içinde olması bakımından benzersizdir. Yumurtalık, stigma, stil ve yumurtalıktan oluşan karpel adı verilen daha büyük bir yapının parçasıdır. Her yumurta, yumurtalık dokusuna funicle adı verilen bir sapla tutturulur. Huninin yumurtalıkla birleştiği noktaya plasenta denir.
Çiçek bir tomurcuktan geliştikçe, archespore adı verilen bir yumurtanın içindeki bir hücre genişleyerek bir embriyo kesesi ana hücresi (EMC) oluşturur. EMC, dört megaspor üretmek için mayoz bölünür . Bu süreçte kromozom sayısı EMC'deki iki setten megasporlardaki bir sete indirilerek megasporlar haploid hale getirilir. Dört megaspordan üçü dejenere olur ve kaybolur, dördüncü ise sekiz haploid hücre üretmek için mitotik olarak üç kez bölünür. Bu hücreler birlikte embriyo kesesi adı verilen dişi gametofitini oluşturur.
Sekiz embriyo kesesi hücresi, iki sinerjide, üç antipodal hücreye, iki kaynaşmış endosperm çekirdeğine ve bir yumurta hücresine farklılaşır. Olgun embriyo kesesi, ovülün dış açıklığında (mikropil) bulunur ve erkek gametofit tarafından verilen sperm hücrelerini almaya hazırdır.

Polen
Erkek gametofit, olgun polen tanesidir. Polen, filamentlerin uzak ucuna tutturulmuş anterlerde üretilir . İplik ve anter birlikte erkek cinsiyet organı olan ercik'i oluşturur. Çiçekler genellikle yaprakların hemen içinde birçok organ üretir. Çiçek olgunlaştıkça, anterdeki hücreler mitotik olarak bölünerek polen ana hücreleri (PMC) üretirler. PMC'ler, tetrad adı verilen dörtlü gruplar halinde haploid mikrosporlar üretmek için mayoz bölünür. Mikrosporlar, gelişen polen tanelerine besin sağlayan tapetum adı verilen tek bir hücre katmanı içinde barındırılır.
Her mikro gözenek, sporopollenin adı verilen bir lipoproteinden oluşan, ekzin adı verilen sert, opak bir dış katman geliştirir . Ekzin, fosil polenlerde bile bir türü tanımlamak için sıklıkla kullanılabilen karakteristik gözeneklere, sırtlara veya çıkıntılara sahiptir. Mikro gözenek, olgun polen tanesinin içinde iki veya üç haploid çekirdek üretmek için mitotik olarak bir veya iki kez bölünür. Çekirdeklerden ikisi, sonunda embriyo kesesinin yumurta ve endosperm çekirdekleriyle kaynaşarak sırasıyla bir embriyo ve endosperm üreten sperm çekirdeği olarak işlev görür.
Bununla birlikte, cinsel füzyonun gerçekleşmesi için polen tanesinin, stilin tepesindeki alıcı bir platform olan stigmaya taşınması gerekir; bu, karpel (ler) in üzerinde uzun bir uzantıdır. Burada nemli yüzey veya kimyasallar polen tanesinin filizlenmesine neden olur. Çimlenme, bir polen tanesinin yüzeyinden bir tüpün büyümesidir. Tüp , içinde su, çözünen maddeler ve herhangi bir hücre duvarı olmayan iki veya üç çekirdek içeren bir pektin kılıfıdır . Polen tüpünün uygun şekilde büyümesi için sulu pektin sentezine yardımcı olabilecek boron gibi besin maddelerinin yanı sıra uygun çözünen konsantrasyonlu çözelti.

Tüpün tepesinde protein sentezinde rol oynayan aktif ribozomlar ve endoplazmik retikulum (hücre organelleri türleri ) bulunur . Pektinaz ve bir glukanaz ( karbonhidratları parçalayan her iki enzim) ) muhtemelen büyüyen tüpün esnekliğini korur ve penetrasyona yardımcı olur. Polen tüpünün tepesi ayrıca ribonükleik asit (RNA) ve ribozomları stilin dokularına salar. Tüp sonunda yumurtalığa ulaşmak için büyür ve burada plasentaya ulaşana kadar hücreler arası boşluklar boyunca hareket edebilir. Kimyasal tanıma yoluyla, polen tüpü büyüme yönünü değiştirir ve plasentadan yumurtaya nüfuz eder. Burada tüp mikropile yakın yatan embriyo kesesine ulaşır ve cinsel döllenme gerçekleşir.
Çift Gübreleme
Çiçekli bitkilerde döllenme, bilinen tüm organizmalar arasında benzersizdir, çünkü bir değil iki hücre çift döllenme adı verilen bir süreçte döllenir. Polen tüpündeki bir sperm çekirdeği embriyo kesesindeki yumurta hücresi ile, diğer sperm çekirdeği diploid endosperm çekirdeği ile birleşir. Döllenmiş yumurta hücresi a, zigot sporofit diploid embriyo geliştirir. Döllenmiş endosperm çekirdeği triploide dönüşür endosperm, embriyo ve fideyi besleyen besleyici bir doku. Çift döllenme sergileyen bilinen diğer tek bitki grubu , çiçeklenmeyen bir tohum bitkisi olan Ephedra cinsindeki Gnetales'tir. Bununla birlikte, bu durumda ikinci döllenme ürünü dejenere olur ve endosperme dönüşmez.
Çifte döllenme, polen tüpü, muhtemelen kalsiyumun kimyasal çekiciliğinin bir sonucu olarak, embriyo kesesindeki iki sinerjid hücreden birine dönüştüğünde başlar. Sinerjide nüfuz ettikten sonra, polen tüpünün tepesi açılır ve iki sperm çekirdeğini ve diğer içerikleri sinerjide bırakır. Sinerjit dejenere olurken, yumurta ve endosperm hücrelerini sararak iki sperm çekirdeğini yakın ve diğer polen tüpünün dışarı atılan içeriğini tutar. Yumurta hücresi daha sonra, zarı parçalanan ve çekirdeğin yumurta çekirdeğine yakın hareket etmesini sağlayan sperm hücresini açar ve yutar. nükleer zarflar daha sonra parçalanır ve iki çekirdek birleşerek zigotun tek diploid çekirdeğini oluşturur. Diğer sperm hücresi, iki endosperm çekirdeği ile birleşerek tek bir triploid hücre oluşturur, birincil endosperm hücresi mitotik olarak endosperm dokusuna bölünür.
Çift döllenme ve endosperm üretimi, fidelerin büyümesini hızlandırarak ve bu savunmasız aşamada hayatta kalmayı artırarak çiçekli bitkilerin büyük ekolojik başarısına katkıda bulunmuş olabilir. Daha hızlı fide gelişimi, çiçek açan bitkilere bazı habitatlarda açık tohumlu bitkilerle rekabette üstünlük sağlamış ve çoğu ılıman ve tropikal bölgelerde çiçekli bitkilerin bolluğuna yol açmış olabilir. Gymnospermler yine de yüksek rakımlarda ve enlemlerde ve Pasifik Kuzeybatı kıyı sekoyaları gibi iğne yapraklı ormanlarda alçak rakımlarda hâkimdir. Bu modellerin nedenleri hala tartışmalı.
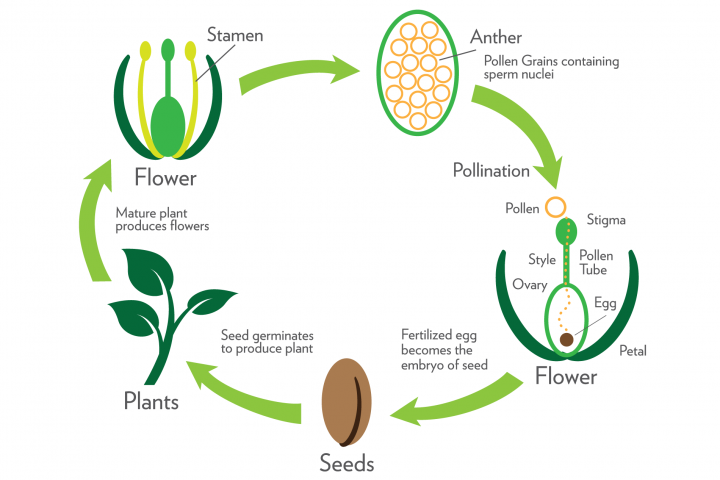
Tohum
Tohum, olgun, döllenmiş ovüldür. Döllenmeden sonra embriyo kesesinin haploid hücreleri parçalanır. Ovülün maternal olarak türetilen diploid hücreleri, tohumun testa veya tohum kabuğu adı verilen sert, suya dayanıklı dış kaplamasına dönüşür. Diploid zigot embriyoya dönüşür ve triploid endosperm hücreleri çoğalarak besin sağlar. Testa genellikle, yumurtanın başlangıçta huniye bağlandığı hilum adı verilen bir yara izini gösterir. Bazı tohumlarda raphe denen testa boyunca uzanan bir çıkıntı, huninin yumurtaya ilk olarak nerede bastırıldığını gösterir. Ovülün mikropili, genellikle tohumun çimlenmesi sırasında suyun geçişine izin veren tohum kabuğunda küçük bir gözenek olarak yaşar.
Bazı türlerde huni, tohum üzerinde aril adı verilen daha büyük bir yapıya dönüşür, bu genellikle parlak renkli, sulu ve aynı zamanda tohumu dağıtabilen hayvanlar tarafından tüketilen şekerler içerir (küçük hindistan cevizi, ararot, oksalis ve teker fasulyesi). Bu, yumurtalıkta oluşan meyveden farklıdır.
Embriyo, kotiledon (lar) , epikotil ve hipokotilden oluşur. Kotiledonlar küçük yapraklara benzer ve genellikle bitkinin ilk fotosentetik organlarıdır. Embriyonun kotiledonların üzerindeki kısmı epikotildir ve aşağıdaki kısım hipokotildir. Epikotil, büyüyen bitkinin sürgününü ve çimlenmeden sonra ilk gerçek yaprakları üreten apikal bir meristemdir . Hipokotil, kökte gelişir. Genellikle hipokotilin ucu olan kök, tohumdan koptuğu için çimlenmenin ilk göstergesidir. Çiçekli bitkiler, monokotiledon veya dikotiledon olarak sınıflandırılır (çoğu artık ekotiledon olarak adlandırılmaktadır) ) embriyoda üretilen kotiledon sayısına göre. Yaygın monokotiledonlar arasında otlar, sazlar, zambaklar, süsen ve orkideler; Yaygın dikotiledonlar arasında ayçiçeği, gül, baklagiller, aslanağzı ve tüm iğne yapraklı olmayan ağaçlar bulunur.
Endosperm, çimlenme sırasında kotiledonları besin kaynağı olarak kullanan birçok baklagillerde olduğu gibi embriyo tarafından tüketilebilir. Diğer türlerde endosperm, gıda olarak kullanıldığında çimlenene kadar devam eder.

Meyve
Çiçekli bir bitkinin meyvesi olgun yumurtalıktır. Tohumlar olgunlaştıkça çevreleyen yumurtalık duvarı, dağılmaya yardımcı olabilecek koruyucu bir yapı oluşturur. Çevreleyen yumurtalık dokusu perikarp olarak adlandırılır ve üç katmandan oluşur. Dıştan içe bu katmanlar ekzokarp, mezokarp ve endokarptır. Ekzokarp genellikle sert ve cilde benzer. Mezokarp genellikle kalın, etli ve tatlıdır. Tohumları çevreleyen endokarp, kayısı gibi etli meyveli çoğu türde olduğu gibi sert ve taşlı olabilir.
Bir meyve, tek bir çiçekte (elma, portakal, kayısı) tek bir olgunlaşmış yumurtalık tarafından üretiliyorsa basit olarak adlandırılır. Bir toplanmış meyve tek çiçek (böğürtlen, ahududu, çilek) tarafından üretilen olgun yumurtalıkların kümesidir. Çoklu meyve, aynı çiçeklenme içinde (ananas, dut, incir) birlikte büyüyen ayrı çiçekler üzerinde birçok olgunlaşmış yumurtalık kümesidir . Basit bir meyve etli veya kuru olabilir. Etli basit bir meyve, dut (üzüm, domates, papaya), pepo (salatalık, karpuz, balkabağı), hesperidium (portakal), sert çekirdekli (kayısı) veya yumuşak çekirdekli (elma) olarak sınıflandırılır.
Kuru basit meyveler, olgunlukta kuru bir perikarpa sahiptir. Tohumları serbest bırakmak için bir dikiş boyunca ayrılabilirler veya ayrılmayabilirler. Bölünmüş kuru meyve, baklagil veya bakla (bezelye, fasulye), silik veya silika (hardal), kapsül (haşhaş, zambak) veya folikül (süt otu, larkspur, columbine) olarak sınıflandırılır. Tohum salmak için ayrılmayan, bağımsız bir kuru meyve, aken (ayçiçeği, düğün çiçeği, çınar), tahıl veya karyops (mısır, buğday, pirinç, arpa gibi çimler), şizokarp (havuç, kereviz, rezene), kanatlı olarak sınıflandırılır. samara (akçaağaç, dişbudak, karaağaç), fındık (meşe palamudu, kestane, fındık) veya utricle (su mercimeği ailesi). Bazı meyve veren cisimler yumurtalık olmayan doku içerir ve bazen psödokarp olarak adlandırılır. Örneğin elma ve armutların tatlı eti,
Her tür meyve veren gövde, içerdikleri tohumları koruma ve dağıtma işlevi görür. Koruma, fiziksel (sert kaplamalar) veya kimyasal (tohum avcılarının kovucuları) olabilir. Tatlı, etli meyveler, meyvelerle birlikte tohumları tüketen ve tohumları gübre görevi görebilecek dışkıda bozulmadan geçiren kuşlar ve memeliler için çekici besindir. Kuru meyveler genellikle, örneğin kanat benzeri yapılar veya kaldırma kuvveti sağlayan kabarık bir pappus yardımıyla tohumların rüzgarla yayılması için uyarlanır. Meyve veren yapıların çeşitliliği, kısmen, farklı meyve boyutu, şekli ve kimyası için seçim yapan ortamdaki dağıtıcı ajanların çeşitliliğini yansıtır.
Tozlaşma ve Tozlayıcılar
Tozlaşma, polenlerin stamenlerden stigmaya doğru hareketidir, burada polen tüpünün çimlenmesi ve büyümesi meydana gelir. Tüm çiçekli bitki türlerinin çoğu (yaklaşık yüzde 96'sı) hermafrodittir (her iki bitki içinde, genellikle her çiçeğin içinde hem cinsel işlevlere sahiptir) ve bu nedenle, bir birey kendi poleni veya başka bir bireyden gelen polenle döllenebilir. Kendi kendine tozlaşma yoluyla üretilen tohum ("kendi kendini dölleyen" tohum) genellikle büyüme, hayatta kalma ve dış melezleme yoluyla üretilen tohuma doğurganlık açısından yetersizdir. tozlaşma ("aşılmış" tohum). Sonuç olarak, çoğu türde, aşılan tohum oranını en üst düzeye çıkarmak için güçlü bir doğal seçilim vardır ("aşma oranı").
Çiçekli bitkiler, çapraz tozlaşma ajanları olarak hayvanları (özellikle böcekler) üstün bir şekilde sömürmeleri açısından tohumlu bitkiler arasında alışılmadık bir durumdur. Böceklerin, kuşların ve memelilerin (özellikle yarasaların) çapraz tozlaşma verimliliği, çiçekli bitkilerin hem bolluğuna hem de çeşitliliğine katkıda bulunmuş olabilir. Başarısız polen ve yumurtalarda daha az enerji ve kaynak israfı nedeniyle bolluk artmış olabilir. Çeşitlilik iki nedenden dolayı artmış olabilir. Birincisi, kuşkusuz böcekler, farklı tozlayıcılara farklı şekillerde farklı ödüller (polen ve nektar) sağlayan ve görünüş (renk yan yana, boyut, şekil) ve koku (tatlı, kokulu) sağlayan çok çeşitli çiçek formlarını seçmişlerdir. İkinci,
Çiçekli bitkiler ilk olarak büyük böcek gruplarının çoğu zaten evrimleştikten sonra ortaya çıksa da, çiçekli bitkiler muhtemelen bu gruplar içindeki birçok yeni türün evrimleşmesine neden oldu. Arılar ve kelebekler gibi bazı yeni böcek grupları, çiçek açan bitkilerden sonra ortaya çıktı, üyeleri ağız kısmı yapıları ve tozlaşma için özel davranış geliştirdiler. Aşırı durumlarda, bir bitki tozlaşma için tamamen bir böcek türüne bağımlıdır ve böcek, besin için tamamen bir bitki türüne bağımlıdır. Böylesine sıkı karşılıklı bağımlılık nadiren meydana gelir, ancak yuccas / yucca güveleri, senita kaktüsleri / senita güveleri ve incir ağaçları / bazı incir eşekarısı ile iyi bir şekilde belgelenmiştir. Her üç böceğin de dişileri çiçeklere yumurta bırakır. ve yavruları daha sonra olgun meyve ve içindekilerle beslenmek için yumurtadan çıkar. Dişiler, başka bir bitkiden polen toplayarak ve onu yumurtalarını tutan çiçeğin stigmasına taşıyarak meyvenin gelişmesini sağlar. Bitkiler, tüketilen bazı meyve ve tohumların küçük bir maliyetiyle üretilen, çaprazlanmış tohumdan büyük ölçüde yararlanır ve böcekler, bitkiler arasındaki kısa mesafelerde polen taşımak için küçük maliyetle larvaları geliştirmek için besin tedarikinden büyük ölçüde yararlanır.
İster biyotik ister abiyotik olsun, tozlaştırıcı ajanlar çiçeğin tüm yönlerine güçlü bir seleksiyon uygulayarak muazzam çiçek çeşitliliğinin evrimleşmesine neden oldu. Bu çeşitlilik, az sayıda karakteristik tozlaşma sendromu olarak damıtılmıştır.
Böcekler tarafından tozlaşma, genellikle beyaz renk, güçlü bir meyve kokusu ve sığ, kase şeklindeki bir çiçek için seçilir. Arılar sarı veya mavi / mor renklendirmeyi, arıyı nektara yönlendiren renk desenlerine sahip bir iniş platformu (genellikle spektrumun morötesi aralığında yansır), iki taraflı simetri ve tatlı bir koku seçerler. Kelebekler sarıdan başka birçok rengi, dibinde nektar bulunan bir korolla (petal) tüpü ve kokusuzluğu seçerler. Tersine güveler gece için seçer güçlü bir koku ve donuk veya beyaz renkte çiçek açmak ve ayrıca tabanında nektarlı bir tüp. Yarasalar gece açan çiçekler için de seçerler, ancak güçlü bir misk kokusu ve bol nektarıyla, kolay erişim için yeşilliklerin çok dışına yerleştirilmiş ve sıkıcı veya beyaz renklidir. Sinek kuşları, kokusuz, bol nektar üretimi ve dibinde nektarlı bir korolla tüpü olmayan kırmızı veya turuncu çiçekleri seçerler. Beslenirken havada asılı kalmayan diğer tozlaşan kuşlar, bol miktarda nektar (tüpler, huniler, fincan şekilleri) içerebilen güçlü tünekleri ve çiçekleri seçerler.
Tozlaştırıcı bir madde olarak rüzgar, renk, koku ve nektar eksikliğini seçer; küçük korolla; geniş bir stigmatik yüzey alanı (genellikle tüylü); bol miktarda üretilen, yüzer polen; ve genellikle dik stamen ve sarkık, stamenleri asılı. Ek olarak, bu sendromların herhangi birinde, üye bitki türlerinin çeşitli evrimsel geçmişlerinden kaynaklanan büyük çiçek çeşitliliği vardır.

Kendinden üreme
Ağaçların, çalıların ve çok yıllık bitkilerin büyük çoğunluğu da dahil olmak üzere, çiçekli bitki türlerinin çoğu öncelikle çaprazlama yoluyla çoğalır. Kendi kendine döllenmeyi önleyen adaptasyonlar arasında kendi kendine uyumsuzluk (kendi kendine polenin genetik olarak tanınması ve engellenmesi) ve dioecy (ayrı erkek ve dişi bireyler) bulunur. Hermafroditlerde kendi kendine tozlaşma olasılığını azaltan adaptasyonlar , anterlerin ve damgaların uzayda (herkogamy) veya zamanda (dikogami) ayrılmasını içerir. Pek çok türde, hem kendi kendine uyumsuzluk hem de cinsiyet organlarının mekansal-zamansal ayrılması meydana gelir.
Bununla birlikte, kendi kendine tohum üretme yeteneği, tozlaşmayı aşmanın zor veya imkansız olduğu durumlarda avantajlıdır. Bunlar, tozlayıcıların nadir veya öngörülemez olduğu zorlu ortamları ve hayatta kalanların genellikle birbirlerinden izole oldukları, düzenli olarak rahatsız edilen zeminleri içerir. Selfing aynı zamanda aşmaktan daha ucuzdur, çünkü selferler hayvanların yardımı olmadan tozlaşabilir ve bu nedenle bol miktarda nektar ve polen ödülü olan büyük, çekici çiçekler üretmeleri gerekmez.
Çoğunlukla kendi kendine yeten türler, değişken veya rahatsız habitatlarda küçük, sıkıcı çiçeklere sahip küçük yıllıklardır. Örneğin, çoğu çöl yıllıkları ve yol kenarındaki yabani otlar selferdir. Outcrossing'den neredeyse tamamen kendi kendine oluşmaya evrimsel geçiş, çiçekli bitkilerde birçok kez meydana geldi.
Outcrossing ve selfing türlerin evrimsel potansiyelleri farklıdır. Outcrosser'lar genellikle genetik olarak daha çeşitlidir ve birçok yeni türün oluştuğu uzun evrimsel dönemler boyunca devam eden soylar üretirler . Bununla birlikte, selfer'lar genetik olarak daha az çeşitlidir ve zararlı mutasyonlar biriktirme eğilimindedir. Genellikle yeni türler geliştirme fırsatı bulamadan nesli tükenir.
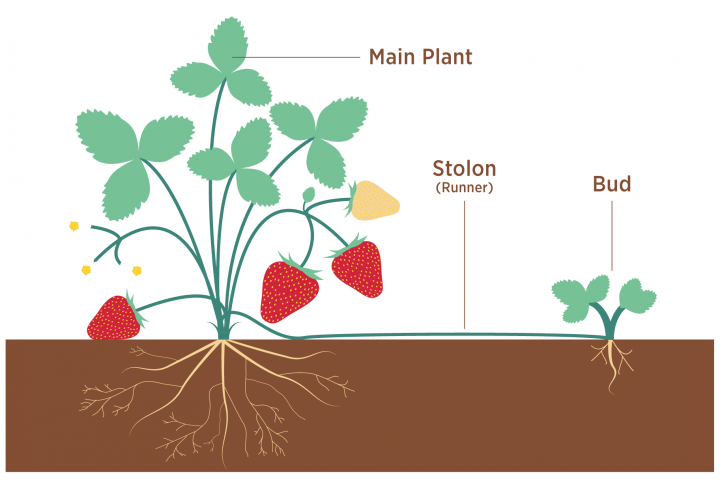
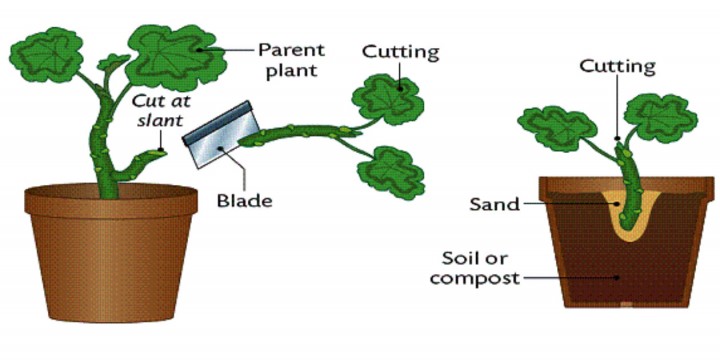
Eşeysiz üreme
Eşeysiz olarak yeni bireyler üretme yeteneği bitkilerde yaygındır. Uygun deneysel koşullar altında, çiçekli bir bitkinin hemen hemen her hücresi tüm bitkiyi yenileyebilir. Doğada yeni bitkiler, uygun bir uyarıcı alan ve ana bitkiden ayrılan yapraklardan, gövdelerden veya köklerden yeniden üretilebilir. Çoğu durumda, bu yeni bitkiler, ebeveynden ayrılmadan önce veya sonra kökler ve filizler üreten tomurcuklara dönüşen farklılaşmamış parankim hücrelerinden ortaya çıkar.
Yer üstü veya yer altı yatay koşuculardan (çilek stolonsu, birçok otun rizomu), yumrulardan (patates, Kudüs enginarı, dahlia), soğanlar (soğan, sarımsak), soğanlar (çiğdem, glayola), sürgündeki soğanlardan yeni bitkiler üretilebilir. (zambak, birçok ot), yapraklardaki parankim hücreleri (Kalanchoe, Afrika menekşesi, yeşim bitkisi) ve çiçeklenme (ok başı). Bitkisel çoğaltma, değerli tarım bitkilerini kesimler, katmanlama ve aşılama yoluyla kopyalamak için ekonomik açıdan önemli bir araçtır. . Vejetatif üreme özellikle suda yaşayan vasküler bitkilerde (örneğin, surfgrass ve eelgrass) yaygındır, bunlardan fragmanlar kopabilir, akıntıda dağılabilir ve yeni bütün bitkiler halinde gelişebilir.
Çiçekli bitkilerin az bir kısmı, yumurta ve sperm füzyonu olmadan tohum üretebilir (partenokarpi veya agamospermi olarak bilinir). Bu, ovüldeki mayoz bölündüğünde ve döllenmeden zigot olarak işlev gören bir diploid yumurta hücresi üretildiğinde meydana gelir. Bilinen örnekler arasında narenciye, karahindiba, hawkweed, düğün çiçeği, böğürtlen / ahududu ve sorbus bulunur. Agamosperm türler, yüksek rakımlarda ve yüksek enlemlerde daha yaygındır ve neredeyse tümü, yakın evrimsel tarihlerinde kromozom sayılarının (tetraploidi) iki katına çıktığını deneyimlemiştir. Bu türler, selfer'larınkine benzer evrimsel avantaj ve dezavantajlar yaşarlar.
Bitki Üreme Stratejilerinin Evrimsel Önemi
En bol ve çeşitli kara bitkileri grubunun - çiçekli bitkilerin - çekici, renkli ve benzersiz özelliklerinin, öncelikle çapraz üremenin verimliliğini ve hızını en üst düzeye çıkarmak için evrimleştiğine inanılmaktadır. Her bir kapalı tohumlu evrim patlaması, polen ve tohumları her zamankinden daha verimli ve çeşitli şekillerde dağıtmak için sömürülen, başta böcekler olmak üzere ilişkili hayvanlarla birlikte evrimsel bir olaydı.
Çiçekli bitki evriminin ilk büyük patlaması, kapalı karpelin radyal olarak simetrik olan gösterişli çiçeklerle birlikte ortaya çıkmasıydı . Kapalı karpel, stilin özel iletken dokusu içindeki kendi polenini tanımak ve bloke etmek yoluyla kendi kendine döllenmeyi önledi. Gösterişli çiçeklere çekilen böcekler, bitkiler arasında polenleri rüzgardan daha az savurgan bir şekilde taşıdılar ve radyal simetri birçok boyut ve şekildeki böcekleri barındırıyordu.
İkinci büyük patlama, arıların evrimleşmesiyle aynı anda birçok bitki grubunda bağımsız olarak gerçekleşen iki taraflı simetrik çiçeklerin ortaya çıkmasıydı . İkili simetri, arıları çiçeklere daha hassas bir şekilde girip çıkmaya zorlayarak daha da verimli outcross polen transferini teşvik etti.
Çiçekli bitki evriminin üçüncü büyük patlaması, kuşların ve kemirgenlerin çeşitliliğiyle çakışan besleyici, etli meyveler ve tohumların ortaya çıkmasıydı. Omurgalıların meyve ve tohum dağıtımı için sömürülmesi, yavruların aynı türden komşu popülasyonlara daha az gelişigüzel taşınmasıyla sonuçlandı (aynı zamanda bir besin kaynağı olarak da ziyaret edildi), böylece yavruların kardeşleri ve ebeveynleri ile aynı soydan çiftleşme şansını düşürdü ve rüzgardan daha fazla güvence sağladı. iyi bir yaşam alanı buldukları akıntılar ve aynı türün ilgisiz çiftleşme partnerleri.
Ahmet TAŞÇI
Kaynaklar:
Ayala, F. J., W. M. Fitch, and M. T. Clegg, eds. Variation and Evolution in Plants and Microorganisms. Washington, DC: National Academy Press, 2000.
Barth, F. G. Insects and Flowers. Princeton, NJ: Princeton University Press, 1991.
Baskin, C. C., and J. M. Baskin. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. San Diego, CA: Academic Press, 1998.
Briggs, D., and S. M. Walters. Plant Variation and Evolution, 3rd ed. New York: Cambridge University Press, 1997.
Chittka, L., and J. D. Thomson, eds. Cognitive Ecology of Pollination: Animal Behavior and Floral Evolution. New York: Cambridge University Press, 2001.
Dickison, W. C. Integrative Plant Anatomy. San Diego, CA: Harcourt/Academic Press, 2000.
Heywood, V. H., ed. Flowering Plants of the World. Oxford: Oxford University Press, 1993.
Marshall, C., and J. Grace. Fruit and Seed Production: Aspects of Development, Environmental Physiology and Ecology (Society for Experimental Biology Seminar, No. 47). New York: Cambridge University Press, 1992.
Proctor, M., P. Yeo, and A. Lack. The Natural History of Pollination. Portland, OR: Timber Press, 1996.
Richards, A. J. Plant Breeding Systems, 2nd ed. Cheltenham, England: Stanley Thornes Publishers Ltd., 1997.
Simpson, B. B., and M. Conner-Orgorzaly. Economic Botany: Plants in Our World, 2nd ed. New York: McGraw-Hill, 1995.
http://www.biologyreference.com/Re-Se/Reproduction-in-Plants.html#ixzz6kSFSALeJ
Foto:Center for Urban Agriculture-doTERRA-Britannica-Class Notes-Britannica-Biologywise-Mühendis Beyinler-UNH Cooperative Extension - University of New Hampshire